2.小脳による運動学習・運動制御機構
小脳は運動制御・運動学習にかかわります。私たちは、長期抑圧等小脳皮質の各機能が動物個体内の行動制御において果たす役割を、遺伝子改変マウスを利用して検討してきています。マウスで運動制御・運動学習能力を定量的に評価するために、反射性眼球運動の計測システムを作製しました(Iwashita et al 2001)。私たちが計測しているのは、前庭動眼反射(VOR)および視運動性眼球運動(OKR)と呼ばれる頭部回転時の視野のブレを補償する反射です(下図参照)。
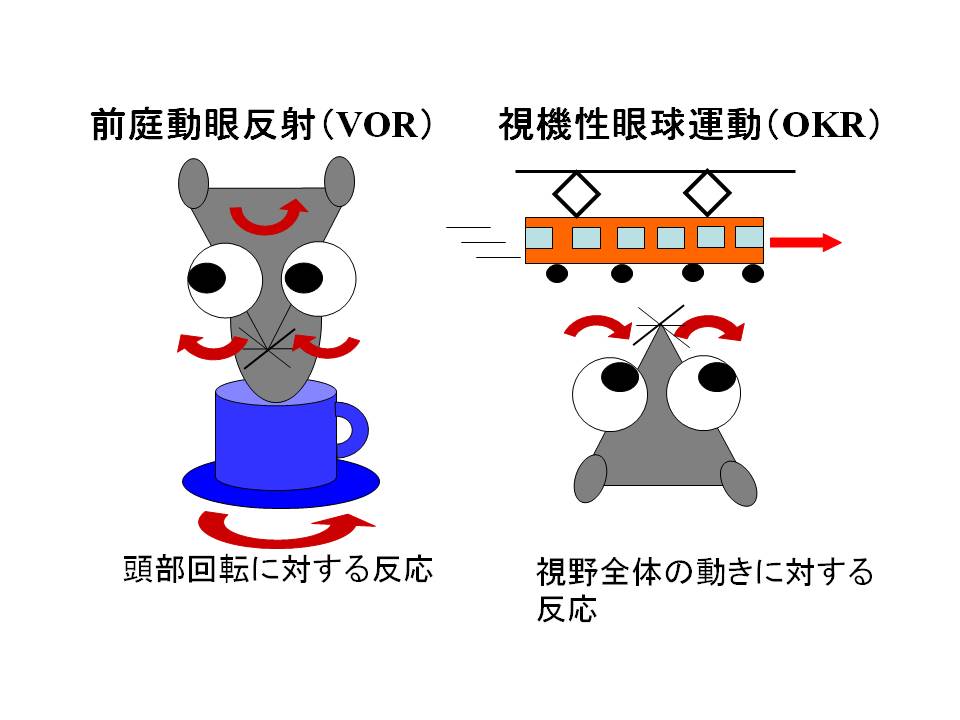
VORは、頭部回転を内耳の半規管が検出し、その情報に基づき眼球を頭部回転と逆方向へ回転させる反射です。一方OKRは、動物の周囲の景色が動いた時に、眼球がその動きにつられて景色の動きと同方向に動く反射です。刺激に対するこれら反射の大きさおよびタイミングを調べることにより、運動制御能力を評価できます。また、これらの反射は状況に応じて適応的に変化し、その変化の様子を調べることにより運動学習能力も評価できます。VORについては、動物を回転させるのと同時に周囲の景色も同方向または逆方向に回転させると、眼球運動の大きさは前者の場合は小さく、後者の場合は大きくなり、視野のブレを小さくするように適応します。またOKRについては、動物周囲の景色の動きがある程度速いと、眼球の動きは視野の動きに十分ついていけないのですが、動物に視野の動きを見続けさせると、眼球の動きがしだいによくなり景色の動きについていけるようになっていく適応が起こります。
私たちは、長期抑圧が起こらないグルタミン酸受容体δ2サブユニット欠損マウスでは、視運動性眼球運動および前庭動眼反射の適応という運動学習が起こらないことを示しました(Katoh et al 2005)。また、δ2サブユニット欠損マウスで不随意の眼球運動が認められ、プルキンエ細胞自体を欠失したラーチャーマウスよりも重篤な運動失調を示すことを見出し、不随意運動発生の神経機構を明らかにしました(Yoshida et al 2004)。さらに、δ2サブユニット欠損マウスで視運動性眼球運動のタイミング遅れが生じることを見出し、そのメカニズムを調べるために、運動中のマウスからのプルキンエ細胞活動記録を行い、δ2サブユニット欠損マウスにおける登上線維入力により引き起こされる複雑スパイクの発火頻度上昇がプルキンエ細胞の活動パターンを変えて、その結果、視運動性眼球運動のタイミングが遅れたことを示唆しました(Yoshida et al 2007)。この研究結果は、神経細胞への異なる種類のシナプス入力強度のバランスが、反射行動タイミングの決定に大きな影響を及ぼすことを示しています。δ2サブユニット欠損マウスでは長期抑圧が不全でしたが、デルフィリン欠損マウスでは長期抑圧が起こりやすくなっていましたので、そのマウスの学習能力を調べ、視運動性眼球運動の適応がより速やかに起こることを明らかにしました(Takeuchi et al 2008)。この結果は、長期抑圧の起こりやすさが運動学習の効率と相関することを示し、一般的にシナプス可塑性の起こりやすさが学習スピードと連関しうることを示唆ています(プレス発表資料1参照)。またプルキンエ細胞上の抑制性シナプスで起こる脱分極依存性増強も運動学習にかかわることを、脱分極依存性増強が起こらないミュータントマウスを用いて明らかにしました(プレス発表参照)。